Cromozomi
Definiție - ce sunt cromozomii?
Alcătuirea genetică a unei celule este stocată sub formă de ADN (acid dezoxiribonucleic) și bazele sale (adenină, timină, guanină și citozină). În toate celulele eucariote (animale, plante, ciuperci) acest lucru este prezent în nucleul celular sub formă de cromozomi. Un cromozom este format dintr-o singură moleculă de ADN coerentă, care este legată de anumite proteine.
Numele de cromozom este derivat din limba greacă și poate fi tradus aproximativ ca „corp de culoare”. Acest nume provine din faptul că foarte devreme în istoria citologiei (1888), oamenii de știință au reușit să le coloreze folosind coloranți de bază speciali și să le identifice la microscopul cu lumină. Cu toate acestea, ele sunt vizibile într-adevăr într-un anumit moment al ciclului celular, mitoza (în celulele germinale, meioza), când cromozomul este deosebit de dens (condensat).

Cum sunt construite cromozomii?
Dacă întreaga helică dublă a ADN a unei celule, adică în jur de 3,4 x 109 perechi de baze, ar fi legată între ele, aceasta ar avea ca rezultat o lungime de peste un metru. Lungimea totală a tuturor cromozomilor adăugați este de numai aproximativ 115 µm. Această diferență de lungime se explică prin structura foarte compactă a cromozomilor, în care ADN-ul este înfășurat sau spiralat de mai multe ori într-un mod foarte specific.
Histonele, o formă specială de proteine, joacă un rol important în acest sens. Există în total 5 histone diferite: H1, H2A, H2B, H3 și H4. Două dintre ultimele patru histone se combină pentru a forma o structură cilindrică, octamerul, în jurul căruia helica dublă se învârte de aproximativ două ori (= super helix). H1 se atașează de această structură pentru a o stabiliza.
Acest complex de ADN, octamer și H1 se numește nucleozom. Mai mulți dintre acești nucleozomi sunt acum „ca un șir de perle” la intervale relativ scurte (10-60 perechi de baze) unul în spatele celuilalt. Secțiunile dintre cromozomi se numesc ADN distanțier. Nucleozomii individuali vin acum în contact din nou prin H1, ceea ce creează o spirală suplimentară și, prin urmare, o compresie.
Catena rezultată, la rândul ei, se află în bucle stabilizate de o coloană vertebrală realizată din proteine acide non-histonice, cunoscute și sub numele de hertonici. Aceste bucle sunt la rândul lor prezente în spirale stabilizate de proteine, ceea ce duce la ultima etapă de compresie. Cu toate acestea, acest grad ridicat de compresie apare doar în contextul diviziunii celulare în timpul mitozei.
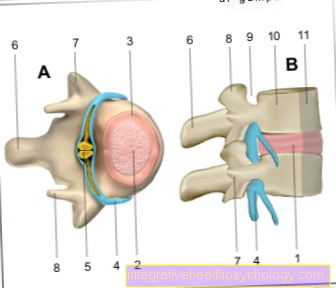
În această fază puteți vedea și forma caracteristică a cromozomilor, care este compusă din două cromatide. Locul în care acestea sunt conectate se numește centromer. Împarte fiecare cromozom metafazic în două brațe scurte și două brațe lungi, numite și brațe p și q.
Dacă centromerul se află aproximativ în mijlocul cromozomului, se numește cromozom metacentric, dacă se află în întregime la unul dintre capetele unui cromozom acrocentric. Cei din mijloc se numesc cromozomi submetacentrici. Aceste diferențe, care pot fi deja văzute la microscopul cu lumină, împreună cu lungimea, permit o clasificare inițială a cromozomilor.
Ce sunt telomerii?
Telomerii sunt capetele cromozomilor cu secvențe repetate (TTAGGG). Acestea nu conțin informații relevante, ci servesc mai degrabă pentru a preveni pierderea unor secțiuni de ADN mai relevante. Cu fiecare diviziune celulară, o parte a cromozomului se pierde prin mecanismul de replicare a ADN-ului.
Deci telomerii sunt, într-un sens, un tampon care întârzie punctul în care celula pierde informații importante prin divizare. Dacă telomerii unei celule au o lungime mai mică de 4.000 de perechi de baze, se inițiază moartea celulară programată (apoptoză). Acest lucru previne răspândirea materialului genetic defect în organism. Câteva celule au telomeraze, adică enzime care sunt capabile să prelungească telomerii din nou.
Pe lângă celulele stem, din care apar toate celelalte celule, acestea sunt celule germinale și anumite celule ale sistemului imunitar. În plus, telomerasele se găsesc și în celulele canceroase, motiv pentru care se vorbește despre imortalizarea unei celule în acest context.
Citiți totul despre subiect aici: Telomeri - Anatomie, funcție și boli
Ce este cromatina?
Cromatina se referă la întregul conținut al unui nucleu celular care poate fi colorat cu o bază. Prin urmare, pe lângă ADN, termenul include și anumite proteine, de exemplu histone și hertonii (vezi structura), precum și anumite fragmente de ARN (hn și snRNA).
În funcție de faza din ciclul celular sau în funcție de activitatea genetică, acest material este disponibil în diferite densități. Forma mai densă se numește heterocromatină. Pentru a face mai ușor de înțeles, s-ar putea deci să o privim ca pe o „formă de stocare” și să facem din nou diferența între heterocromatina constitutivă și facultativă.
Heterocromatina constitutivă este cea mai densă formă, care este prezentă în toate fazele ciclului celular în cel mai înalt nivel de condensare. Acesta reprezintă aproximativ 6,5% din genomul uman și este situat în principal în apropierea centromerilor și a capetelor brațelor cromozomiale (telomeri) într-o mică măsură, dar și în alte locuri (în principal cromozomii 1, 9, 16, 19 și Y) . În plus, cea mai mare parte a heterocromatinei constitutive se află lângă membrana nucleară, adică la marginile nucleului celular. Spațiul din mijloc este rezervat cromatinei active, euchromatinei.
Heterocromatina facultativă este puțin mai densă și poate fi activată și dezactivată după cum este necesar sau în funcție de stadiul de dezvoltare. Un bun exemplu în acest sens este al doilea cromozom X din cariotipurile feminine. Deoarece un cromozom X este practic suficient pentru ca celula să supraviețuiască, așa cum este în cele din urmă suficient pentru bărbați, unul dintre cei doi este dezactivat în faza embrionară. cromozomul X dezactivat este cunoscut sub numele de corp Barr.
Numai în timpul diviziunii celulare, în contextul mitozei, se condensează complet, prin care atinge cea mai mare compresie în metafază. Cu toate acestea, din moment ce diferitele gene sunt citite diferit adesea - la urma urmei, nu fiecare proteină este necesară în aceeași cantitate tot timpul - aici se face o distincție între euchromatină activă și inactivă.
Citiți mai multe despre acest sub: Cromatină
Cromozomi haploizi
Haploid (grecesc haploos = unic) înseamnă că toți cromozomii unei celule sunt prezenți individual, adică nu în perechi (diploide), așa cum se întâmplă de obicei. Aceasta este starea naturală a tuturor ovulelor și spermatozoizilor, în care cele două cromatide identice nu sunt inițial separate în cursul primei meioze, ci în schimb toate perechile de cromozomi sunt separate mai întâi.
Drept urmare, după prima meioză, celulele fiice la om au doar 23 în loc de 46 de cromozomi obișnuiți, ceea ce corespunde cu jumătate din setul haploid de cromozomi. Deoarece aceste celule fiice au încă o copie identică a fiecărui cromozom format din 2 cromozomi, este necesară a doua meioză, în care cele două cromatide sunt separate una de cealaltă.
Cromozomi politenici
Un cromozom politen este un cromozom format dintr-un număr mare de cromatide identice genetic. Deoarece astfel de cromozomi sunt ușor de văzut chiar și sub o mărire mai mică, uneori sunt denumiți cromozomi gigantici. Condiția prealabilă pentru aceasta este endoreplicarea, în care cromozomii din nucleul celulei sunt înmulțiți de mai multe ori fără ca diviziunea celulară să aibă loc.
Care sunt funcțiile cromozomilor?
Cromozomul ca unitate organizațională a genomului nostru servește în primul rând pentru a se asigura că genomul dublat este distribuit uniform între celulele fiice în timpul diviziunii celulare. Pentru a face acest lucru, merită să aruncăm o privire mai atentă asupra mecanismelor de diviziune celulară sau a ciclului celular:
Celula petrece majoritatea ciclului celular în interfață, ceea ce înseamnă întreaga perioadă de timp în care celula nu este pe cale să se împartă imediat. La rândul său, aceasta este împărțită în faze G1, S și G2.
Faza G1 (G ca în decalaj, adică decalaj) urmează imediat diviziunea celulară. Aici celula crește din nou în dimensiune și îndeplinește funcții metabolice generale.
De aici poate trece și la faza G0. Aceasta înseamnă că se schimbă într-o etapă care nu mai este capabilă de divizare și, în cazuri normale, se schimbă foarte mult pentru a îndeplini o funcție foarte specifică (diferențierea celulară). Pentru a îndeplini aceste sarcini, genele foarte specifice sunt citite mai intens, altele mai puțin sau deloc.
Dacă un segment de ADN nu este necesar pentru o lungă perioadă de timp, acesta este adesea localizat în părțile cromozomilor care au fost dens ambalate de mult timp (vezi cromatina). Pe de o parte, aceasta are scopul de a economisi spațiu, dar pe lângă celelalte mecanisme de reglare a genelor, este și o protecție suplimentară împotriva citirii accidentale. Cu toate acestea, s-a observat, de asemenea, că, în condiții foarte specifice, celulele diferențiate din faza G0 pot intra din nou în ciclu.
Faza G1 este urmată de faza S, adică faza în care ADN-ul nou este sintetizat (replicarea ADN). Aici, întregul ADN trebuie să fie în cea mai slabă formă, adică toți cromozomii sunt complet neînfășurați (vezi structura).
La sfârșitul fazei de sinteză, întregul material genetic este prezent în duplicat în celulă. Deoarece copia este încă atașată la cromozomul original prin centromer (vezi structura), nu se vorbește despre o duplicare a cromozomilor.
Fiecare cromozom este acum format din două cromatide în loc de una, astfel încât să poată lua mai târziu forma X caracteristică în timpul mitozei (strict vorbind, forma X se aplică doar cromozomilor metacentrici). În faza G2 ulterioară, are loc pregătirea imediată pentru divizarea celulară. Aceasta include, de asemenea, o verificare detaliată a erorilor de replicare și a rupturilor de șiruri, care pot fi reparate dacă este necesar.
În principiu există două tipuri de diviziune celulară: mitoza și meioza. Cu excepția celulelor germinale, toate celulele unui organism apar prin mitoză, a cărei sarcină este formarea a două celule fiice identice genetic.
Meioza, pe de altă parte, are scopul de a genera celule diferite din punct de vedere genetic:
Într-un prim pas, cromozomii corespunzători (omologi) dar nu identici sunt împărțiți. Numai în etapa următoare sunt cromozomii, care constau din două cromatide identice, separate și distribuite din nou la două celule fiice fiecare, astfel încât, în final, patru celule germinale cu material genetic diferit provin dintr-o celulă precursoră.
Forma și structura cromozomilor sunt esențiale pentru ambele mecanisme: „fire proteice” speciale, așa-numitul aparat fus, se atașează la cromozomii foarte condensați și trag cromozomii într-un proces reglat fin din planul mediu (plan ecuatorial) către polii opuși ai celulei din jurul unuia pentru a asigura o distribuție uniformă. Chiar și mici modificări ale microstructurii cromozomilor pot avea consecințe grave aici.
La toate mamiferele, raportul dintre cromozomii sexuali X și Y determină, de asemenea, sexul descendenților. Practic, totul depinde dacă spermatozoizii care se unesc cu celula ovulă au un cromozom X sau Y. Deoarece ambele forme de spermă sunt întotdeauna produse exact în aceeași măsură, probabilitatea este întotdeauna echilibrată pentru ambele sexe. Acest sistem aleatoriu garantează o distribuție mai uniformă a genului decât ar fi cazul, de exemplu, cu factori de mediu precum temperatura.
Aflați mai multe despre subiect: Diviziunea nucleului celular
Cum se transmite machiajul genetic prin cromozomi?
Astăzi știm că trăsăturile sunt moștenite prin gene care sunt stocate în celule sub formă de ADN. Acestea sunt la rândul lor împărțite în 46 de cromozomi, pe care sunt distribuite cele 25.000-30000 de gene umane.
Pe lângă proprietatea însăși, care se numește fenotip, există și echivalentul genetic, care se numește genotip. Locul în care se află o genă pe un cromozom se numește locus. Deoarece oamenii au dublu fiecare cromozom, fiecare genă apare și de două ori. Singura excepție de la aceasta sunt genele cromozomiale X la bărbați, deoarece cromozomul Y conține doar o fracțiune din informațiile genetice găsite pe cromozomul X.
Diferite gene care se află pe același locus se numesc alele. Adesea există mai mult de două alele diferite la un locus. Se vorbește apoi de polimorfism. O astfel de alelă poate fi pur și simplu o variantă inofensivă (variantă normală), dar și mutații patologice care pot fi declanșatorul unei boli ereditare.
Dacă mutația unei singure gene este suficientă pentru a schimba fenotipul, se vorbește despre moștenire monogenă sau mendeliană. Cu toate acestea, multe dintre trăsăturile moștenite sunt moștenite prin mai multe gene care interacționează și, prin urmare, sunt mult mai dificil de studiat.
Întrucât mama și tată transmit fiecare una dintre cele două gene către copil într-o moștenire mendeliană, există întotdeauna patru combinații posibile în generația următoare, prin care acestea pot fi, de asemenea, aceleași în raport cu o proprietate. Dacă ambele alele ale unui individ au același efect asupra fenotipului, individul este homozigot în raport cu această caracteristică, iar caracteristica este exprimată în mod corespunzător.
Heterozigoții au două alele diferite care pot interacționa între ele în moduri diferite: Dacă o alelă este dominantă asupra alteia, aceasta își suprimă complet expresia și trăsătura dominantă devine vizibilă în fenotip. Alela suprimată se numește recesivă.
În cazul unei moșteniri codominante, ambele alele se pot exprima neafectate una de cealaltă, în timp ce în cazul unei moșteniri intermediare există un amestec de ambele caracteristici. Un bun exemplu în acest sens este sistemul de grupe sanguine AB0, în care A și B sunt co-dominante între ele, dar 0 sunt dominante unul față de celălalt.
Care este setul normal de cromozomi la om?
Celulele umane au 22 de perechi de cromozomi (autozomi) independenți de sex și doi cromozomi sexuali (gonozomi), deci un total de 46 de cromozomi alcătuiesc un set de cromozomi.
Autozomii vin de obicei în perechi. Cromozomii unei perechi sunt similare ca formă și secvență de gene și, prin urmare, sunt denumiți omologi. Cei doi cromozomi X la femei sunt, de asemenea, omologi, în timp ce bărbații au un cromozom X și un Y. Acestea diferă prin forma și numărul de gene prezente în așa fel încât nu se mai poate vorbi de omologie.
Celulele germinale, adică celulele ovule și spermatozoizii, au doar jumătate din setul de cromozomi datorită meiozei, și anume 22 de autozomi individuali și câte un gonosom. Deoarece celulele germinale se fuzionează în timpul fertilizării și uneori schimbă segmente întregi (încrucișare), se creează o nouă combinație de cromozomi (recombinare). Toți cromozomii împreună sunt numiți cariotip, care cu câteva excepții (a se vedea aberațiile cromozomiale) este identic la toți indivizii de același sex.
Aici puteți afla totul despre subiect: Mitoza - Pur și simplu explicată!
De ce există întotdeauna perechi de cromozomi?
Practic, la această întrebare se poate răspunde cu o singură propoziție: Pentru că s-a dovedit a fi benefică.Prezența perechilor de cromozomi și principiul recombinării sunt esențiale pentru moștenire în ceea ce privește reproducerea sexuală. În acest fel, un individ complet nou poate ieși din materialul genetic al a doi indivizi din întâmplare.
Acest sistem mărește enorm varietatea proprietăților din cadrul unei specii și se asigură că se poate adapta la condițiile de mediu modificate mult mai rapid și mai flexibil decât ar fi posibil doar prin mutație și selecție.
Setul dublu de cromozomi are, de asemenea, un efect protector: dacă o mutație a unei gene ar duce la eșecul funcției, există încă un fel de „copie de rezervă” în al doilea cromozom. Acest lucru nu este întotdeauna suficient pentru ca organismul să compenseze defecțiunile, mai ales dacă alela mutantă este dominantă, dar crește șansele apariției acesteia. În plus, în acest fel mutația nu este transmisă automat tuturor descendenților, ceea ce la rândul său protejează specia de mutații excesiv de radicale.
Ce este o mutație cromozomială?
Defectele genetice pot apărea din radiații ionizante (de exemplu, raze X), substanțe chimice (de exemplu, benzopiren în fumul de țigară), anumite virusuri (de exemplu, virusuri HP) sau, cu o probabilitate redusă, pot apărea, de asemenea, pur din întâmplare. Există adesea mai mulți factori implicați în dezvoltarea acestuia. În principiu, astfel de modificări pot apărea în toate țesuturile corpului, dar din motive practice analiza este de obicei limitată la limfocite (un tip special de celule imune), fibroblaste (celule ale țesutului conjunctiv) și celule ale măduvei osoase.
O mutație cromozomială este o schimbare structurală majoră a cromozomilor individuali. Absența sau adăugarea de cromozomi întregi, pe de altă parte, ar fi o mutație a genomului sau a ploidiei, în timp ce termenul mutație genică se referă la modificări relativ mici în cadrul unei gene. Termenul aberație cromozomială (latină aberrare = a devia) este oarecum mai larg și include toate modificările care pot fi detectate cu microscopul cu lumină.
Mutațiile pot avea efecte foarte diferite:
- Mutațiile silențioase, adică mutațiile în care schimbarea nu are niciun efect asupra individului sau descendenților lor, sunt destul de atipice pentru aberațiile cromozomiale și se găsesc mai des în zona mutațiilor genetice sau punctuale.
- Se vorbește despre o mutație de pierdere a funcției atunci când mutația are ca rezultat o proteină greșită și, prin urmare, fără funcție sau deloc proteină.
- Așa-numitele mutații de câștig de funcție modifică tipul de efect sau cantitatea de proteine produse în așa fel încât să apară efecte complet noi. Pe de o parte, acesta este un mecanism crucial pentru evoluție și, prin urmare, pentru supraviețuirea unei specii sau apariția unor specii noi, dar pe de altă parte, ca și în cazul cromozomului Philadelphia, poate contribui, de asemenea, decisiv la dezvoltarea celulelor canceroase.
Cele mai cunoscute dintre diferitele forme de aberații cromozomiale sunt probabil aberațiile numerice, în care cromozomii individuali sunt prezenți o singură dată (monozomie) sau chiar triplu (trisomie).
Dacă acest lucru se aplică doar unui singur cromozom, se numește aneuploidie, iar întregul set de cromozomi este afectat de poliploidie (tri- și tetraploidie). În cele mai multe cazuri, această distribuție neadecvată apare în cursul dezvoltării celulelor germinale prin nedepărtarea (nedisjunctiune) a cromozomilor în timpul diviziunii celulare (meioză). Acest lucru duce la o distribuție inegală a cromozomilor pe celulele fiice și, astfel, la aberații numerice la copil.
Monosomiile cromozomilor non-sexuali (= autozomi) sunt incompatibile cu viața și, prin urmare, nu apar la copiii vii. Cu excepția trisomiilor 13, 18 și 21, trisomiile autozomale conduc aproape întotdeauna la avorturi spontane.
În orice caz, spre deosebire de aberațiile cromozomilor sexuali, care pot fi, de asemenea, puțin vizibile, există întotdeauna simptome clinice grave și, de regulă, anomalii externe mai mult sau mai puțin pronunțate (dismorfisme).
O astfel de distribuție defectuoasă poate apărea și mai târziu în viață, cu diviziune celulară mitotică (toate celulele, cu excepția celulelor germinale). Deoarece există celule neschimbate în plus față de celulele afectate, se vorbește despre un mozaic somatic. Prin somatic (soma grecesc = corp) se înțeleg toate celulele care nu sunt celule germinale. Deoarece doar o mică parte a celulelor corpului este afectată, simptomele sunt de obicei mult mai ușoare. Prin urmare, tipurile de mozaic rămân deseori nedetectate mult timp.
Aici puteți afla totul despre subiect: Mutația cromozomială
Ce este aberația cromozomială?
Aberația structurală a cromozomului corespunde practic definiției mutației cromozomiale (vezi mai sus). Dacă cantitatea de material genetic rămâne aceeași și este distribuită pur și simplu diferit, se vorbește despre o aberație echilibrată.
Acest lucru se face adesea prin translocare, adică prin transferul unui segment cromozomial către alt cromozom. Dacă este un schimb între doi cromozomi, se vorbește despre translocație reciprocă. Deoarece doar aproximativ 2% din genom este necesar pentru a produce proteine, probabilitatea este foarte mică ca o astfel de genă să fie la punctul de rupere și, prin urmare, își pierde funcția sau este afectată în ea. Prin urmare, o astfel de aberație echilibrată trece adesea neobservată și se transmite de-a lungul mai multor generații.
Cu toate acestea, acest lucru poate duce la o distribuție defectuoasă a cromozomilor în timpul dezvoltării celulelor germinale, ceea ce poate duce la infertilitate, avort spontan sau descendenți cu o aberație dezechilibrată.
O aberație dezechilibrată poate apărea, de asemenea, în mod spontan, adică fără antecedente familiale. Probabilitatea ca un copil să se nască viu cu aberație dezechilibrată depinde în mare măsură de cromozomii afectați și variază între 0 și 60%. Acest lucru duce la pierderea (= ștergerea) sau duplicarea (= duplicarea) unui segment de cromozom. În acest context, se vorbește și despre mono- și trisomii parțiale.
În unele cazuri, acestea apar împreună în două regiuni diferite, monosomia parțială fiind de obicei mai decisivă pentru apariția simptomelor clinice. Acestea sunt exemple proeminente de ștergere Sindromul Scream de pisică și Sindromul Wolf-Hirschhorn.
Se vorbește despre o microdelecție atunci când schimbarea nu mai poate fi determinată cu microscopul cu lumină, adică atunci când este vorba despre pierderea uneia sau a câtorva gene. Acest fenomen este considerat a fi cauza sindromului Prader-Willi și a sindromului Angelman și este strâns legat de dezvoltarea retionoblastomului.
Translocarea Robertson este un caz special:
Doi cromozomi acrocentrici (13, 14, 15, 21, 22) se unesc la centromer și formează un singur cromozom după ce brațele scurte au fost pierdute (vezi structura). Deși rezultă un număr redus de cromozomi, aceasta este denumită o aberație echilibrată, deoarece pierderea brațelor scurte din acești cromozomi poate fi ușor compensată. Și aici, efectele sunt adesea vizibile numai în generațiile următoare, deoarece există o probabilitate foarte mare de avorturi spontane sau de copii în viață cu o trisomie.
Dacă există două pauze într-un cromozom, se poate întâmpla ca segmentul intermediar să fie rotit cu 180 ° și încorporat în cromozom. Acest proces, cunoscut sub numele de inversiune, este dezechilibrat numai dacă punctul de rupere se află într-o genă activă (2% din totalul materialului genetic). În funcție de faptul dacă centromerul se află în interiorul sau în afara segmentului inversat, este o inversare peri- sau paracentrică. Aceste modificări pot contribui, de asemenea, la distribuția inegală a materialului genetic pe celulele germinale.
În inversiunea paracentrică, în care centromerul nu se află în segmentul inversat, pot apărea și celule germinale cu doi sau deloc centromer. Ca rezultat, cromozomul corespunzător se pierde în primele diviziuni celulare, ceea ce duce aproape sigur la un avort spontan.
Inserarea este instalarea unui fragment de cromozom în altă parte. Și aici, descendenții sunt afectați în primul rând într-un mod similar. Un cromozom inelar poate apărea în special după ștergerea pieselor finale. Tipul și dimensiunea secvențelor sunt decisive pentru severitatea simptomelor. În plus, acest lucru poate duce la distribuții incorecte și, prin urmare, la tipuri de mozaic în celulele corpului.
Dacă cromozomul metafazic se separă incorect în timpul diviziunii celulare, pot rezulta izocromozomi. Aceștia sunt doi aceiași cromozomi care constau doar din brațe lungi sau doar scurte. În cazul cromozomului X, acesta se poate manifesta ca un sindrom Ulrich-Turner (monozomia X).
Citiți mai multe informații despre acest subiect: Aberație cromozomială
Trisomia 21
Trisomia 21, mai cunoscută sub numele de sindrom Down, este probabil cea mai frecventă aberație numerică cromozomială în rândul nașterilor vii, bărbații fiind afectați puțin mai des (1,3: 1).
Probabilitatea apariției trisomiei 21 depinde de diverși factori demografici, cum ar fi vârsta medie la naștere a mamelor și variază ușor de la o regiune la alta.
95% din trisomia 21 apare ca urmare a unei erori de diviziune în contextul meiozei (diviziunea celulelor germinale), și anume nedisjunctie, adică eșecul de a separa cromatidele surori.
Acestea sunt cunoscute sub numele de trisomii libere și apar 90% la nivelul mamei, 5% la paterne și încă 5% la genomul embrionar.
Un alt 3% rezultă din translocații dezechilibrate fie pe cromozomul 14, fie ca 21; 21 translocație, creând un cromozom normal și un dublu 21. Restul de 2% sunt tipuri de mozaic în care trisomia nu a apărut în celulele germinale și, prin urmare, nu afectează toate celulele corpului. Tipurile de mozaic sunt adesea atât de ușoare încât pot rămâne complet nedetectate pentru o lungă perioadă de timp.
În orice caz, ar trebui efectuată o examinare cromozomială pentru a distinge trisomia liberă simptomatic identică de trisomia de translocație posibil moștenită. Apoi poate urma o istorie familială a generațiilor anterioare.
Te interesează acest subiect? Citiți următorul articol despre asta: Trisomia 21
Trisomia 13
Trisomia 13 sau sindromul Patau are o frecvență de 1: 5000 și este mult mai rar decât sindromul Down. Cauzele (trisomii libere, translocații și tipuri de mozaic) și distribuția procentuală a acestora sunt în mare parte identice.
În teorie, aproape toate cazurile ar putea fi diagnosticate prenatal folosind ultrasunete sau testul PAPP-A. Deoarece testul PAPP-A nu face parte neapărat din examinările de rutină, aproximativ 80% din cazuri în Europa Centrală sunt diagnosticate înainte de naștere.
Un reziduu de creștere, o buză și palatul fisurat bilateral și ochi neobișnuit de mici (microftalmie) pot fi deja văzute în ultrasunete. În plus, sunt de obicei prezente malformații ale creierului anterior și ale feței de diferite grade de severitate (holoprosencefalie).
În timp ce în forma lobară emisferele cerebrale sunt aproape complet separate și se creează ventriculii laterali, în forma semi-lobară adesea doar partea din spate a creierului este separată și ventriculii laterali lipsesc. În forma cea mai severă, forma alobară, nu există o separare a emisferelor cerebrale.
Sugarii cu formă semi- sau alobar mor de obicei imediat după naștere. După o lună, rata mortalității este de aproximativ 50% din nașterile vii. Până la vârsta de 5 ani, rata mortalității din trisomia 13 crește la 90%. Datorită malformațiilor din creier, în majoritatea cazurilor persoanele bolnave rămân la pat pentru toată viața și nu pot vorbi, motiv pentru care sunt dependenți de îngrijirea deplină. În plus, pot exista și manifestări fizice de anvergură ale Trismoie 13.
Citiți mai multe despre acest subiect la: Trisomia 13 la copilul nenăscut
Trisomia 16
Practic, trisomia 16 este cea mai frecventă trisomie (aproximativ 32% din toate trisomiile), dar copiii vii cu trisomia 16 sunt foarte rare. În general, nașterile vii apar doar în trisomii parțiale sau tipuri de mozaic. Cu toate acestea, printre trisomii este cel mai frecvent responsabil pentru nașteri mortale: 32 din 100 avorturi spontane datorate aberațiilor cromozomiale pot fi urmărite înapoi la această formă de trisomie.
Prin urmare, au fost documentate în principal caracteristici identificabile prenatale, adică prenatale. De remarcat aici sunt diverse defecte cardiace, creșterea încetinită, o singură arteră ombilicală (altfel dublă) și o transparență crescută a gâtului, care se explică prin acumularea de lichide datorită sistemului limfatic care nu este încă complet dezvoltat și a elasticității crescute a pielii în această zonă. În plus, hernia ombilicală fiziologică, adică deplasarea temporară a unei părți mari a intestinului prin buric către exterior, adesea nu regresează corespunzător, ceea ce este cunoscut sub numele de omfalocel sau ruptura cordonului ombilical.
O contractură de flexie cu degetele încrucișate poate fi, de asemenea, adesea detectată la ultrasunete. La puținele nașteri vii, se observă hipotensiunea musculară generalizată, adică slăbiciunea musculară generală. Acest lucru duce la slăbiciunea consumului de alcool și poate asigura că sugarul trebuie hrănit artificial. De multe ori apare și brazda cu patru degete, care este atât de caracteristică trisomiilor. Și aici, frecvența apariției trisomiei este direct legată de vârsta mamei.
Trisomia 18
Sindromul Edwards, adică trisomia 18, apare cu o frecvență de 1: 3000. În cazul diagnosticului prenatal, este la fel ca și în cazul sindromului Patau: și aici, aceleași examinări ar permite tuturor pacienților să fie găsiți complet înainte de naștere. Cauzele și distribuția lor pot fi comparate cu alte trisomii (vezi trisomia 21).
În plus, trisomiile parțiale apar în trisomia 18, care, la fel ca tipurile de mozaic, duc la cursuri clinice mult mai ușoare. Dismorfismele asociate sunt, de asemenea, extrem de caracteristice pentru sindromul Edwards: La naștere, pacienții au o greutate corporală foarte redusă de 2 kg (normal: 2,8-4,2 kg), o frunte largă în retragere, o jumătate inferioară a feței în general subdezvoltată, cu o gură mică deschidere, fantele înguste ale pleoapelor și urechile rotite înapoi, schimbate de formă (urechea faunului). Partea din spate a capului, care este neobișnuit de puternic dezvoltată pentru un nou-născut, este, de asemenea, vizibilă. Coaste sunt neobișnuit de înguste și fragile. Nou-născuții au, de asemenea, o tensiune permanentă (tonus) a întregii musculaturi, care, totuși, regresează la supraviețuitori după primele câteva săptămâni.
O altă trăsătură caracteristică este încrucișarea celui de-al doilea și al cincilea deget peste cel de-al treilea și al patrulea cu numărul total de degete ciocănite, în timp ce picioarele sunt neobișnuit de lungi (trecute), au un toc deosebit de pronunțat, unghiile degetelor de la picioare și un deget mare .
Malformațiile grave ale organelor sunt frecvente și apar de obicei în combinație: defecte ale inimii și rinichilor, defectarea (malrotarea) intestinului, aderențe ale peritoneului (comuna mesenterium), o ocluzie a esofagului (atrezie esofagiană) și multe altele.
Din cauza acestor malformații, rata mortalității este de aproximativ 50% în primele 4 zile, doar aproximativ 5-10% trăiesc până la vârsta de peste un an. Supraviețuirea până la vârsta adultă este excepția absolută. În orice caz, o dizabilitate intelectuală este foarte pronunțată și nu poate vorbi, este așezată la pat și incontinentă, deci complet dependentă de ajutorul din exterior.
Pentru informații mai detaliate despre trisomia 18, vă rugăm să citiți și articolul nostru detaliat pe această temă:
- Trisomia 18 (Sindromul Edwards)
- Trisomia 18 la copilul nenăscut
Trisomia X
Trisomia X este cea mai vizibilă formă de aberație numerică cromozomială, aspectul celor afectați, care sunt în mod logic toți de sex feminin, nu diferă foarte mult de celelalte femei. Unele se remarcă pentru că sunt deosebit de înalte și au trăsături faciale oarecum „plinuțe”. Dezvoltarea mentală poate fi, de asemenea, în mare măsură normală, variind de la limita normală la dizabilitatea mentală ușoară.
Cu toate acestea, acest deficit de inteligență este oarecum mai grav decât în cazul celorlalte trisomii ale cromozomilor sexuali (XXY și XYY). Cu o frecvență de 1: 1000, de fapt, nu este atât de rar, dar întrucât trisomia nu este de obicei asociată cu simptome semnificative clinic, majoritatea femeilor cu boală nu vor fi diagnosticate niciodată pentru întreaga lor viață.
Purtătorii sunt descoperiți în mare parte din întâmplare în timpul unui control familial sau în timpul diagnosticului prenatal.Fertilitatea poate fi ușor redusă, iar rata aberațiilor cromozomiale sexuale în generația următoare poate fi ușor crescută, astfel încât consilierea genetică este recomandată dacă doriți să aveți copii.
Ca și în cazul celorlalte trisomii, trisomia X se dezvoltă cel mai adesea ca o trisomie liberă, adică datorită lipsei de diviziune (nedisjunctie) a cromatidelor surori. Și aici apare de obicei în timpul maturării celulelor ovale materne, deși probabilitatea crește odată cu vârsta.
Sindromul X fragil
Sindromul X fragil sau sindromul Martin Bell este preferat la bărbați, deoarece aceștia au doar un cromozom X și, prin urmare, sunt mai afectați de schimbare.
Apare cu o frecvență de 1: 1250 în rândul nașterilor masculine vii într-un an, făcându-l cea mai comună formă de întârziere mintală nespecifică, adică toate handicapurile mentale care nu pot fi descrise de un sindrom special cu semne tipice.
Sindromul X fragil poate apărea de obicei la fete într-o formă oarecum mai slabă, care se datorează inactivării accidentale a unuia dintre cromozomii X. Cu cât este mai mare proporția cromozomului X sănătos oprit, cu atât simptomele sunt mai puternice.
De cele mai multe ori, totuși, femeile sunt purtătoare ale premutației, care încă nu produce simptome clinice, dar crește masiv probabilitatea de mutație completă la fiii lor. În cazuri foarte rare, bărbații pot fi, de asemenea, purtătorii premutației, pe care îi pot transmite apoi numai fiicelor care, cu toate acestea, sunt de obicei sănătoase din punct de vedere clinic (paradoxul Sherman).
Sindromul este declanșat de un număr extrem de crescut de triplete CGG (o anumită secvență de bază) în gena FMR (fragil-site-mental-retardation); în loc de 10-50 de exemplare, premutația 50-200, când este complet dezvoltată 200- 2000 exemplare.
La microscopul cu lumină, acest lucru pare a fi o ruptură în brațul lung, ceea ce a dat numele sindromului. Acest lucru duce la dezactivarea genei afectate, care la rândul său provoacă simptomele.
Persoanele afectate prezintă o dezvoltare încetinită a vorbirii și a mișcării și pot prezenta probleme comportamentale care pot merge în direcția hiperactivității, dar și a autismului. Anomaliile pur externe (semne de dismorfism) sunt o față lungă cu bărbie proeminentă și urechi proeminente. Odată cu pubertatea, testiculele sunt adesea mărite foarte mult (macroorchidia), iar trăsăturile feței devin mai grosiere. Există o ușoară acumulare de anomalii psihologice și o menopauză deosebit de timpurie în rândul femeilor purtătoare ale premutației.
Ce este o analiză cromozomială?
Analiza cromozomială este un proces în citogenetică cu care pot fi detectate aberații numerice sau structurale ale cromozomilor.
O astfel de analiză ar fi utilizată, de exemplu, dacă se suspectează imediat un sindrom cromozomial, adică în cazul malformațiilor (dismorfismelor) sau al dizabilităților intelectuale (întârziere), dar și în cazul infertilității, avorturilor regulate (avorturi) și, de asemenea, cu anumite tipuri de cancer (de exemplu limfoame sau leucemie).
Acest lucru necesită de obicei limfocite, un tip special de celule imune care se obține din sângele pacientului. Deoarece numai o cantitate relativ mică poate fi obținută în acest mod, celulele sunt stimulate să se împartă cu fitohemaglutinina, iar limfocitele pot fi apoi cultivate în laborator.
În unele cazuri, se prelevează probe (biopsii) din piele sau din măduva spinării, cu o procedură similară. Scopul este de a obține cât mai mult material ADN care este în prezent în mijlocul diviziunii celulare. În metafază, toți cromozomii sunt dispuși într-un nivel aproximativ în mijlocul celulei, pentru a fi atrași pe laturile opuse (polii) celulei în etapa următoare, anafaza.
În acest moment, cromozomii sunt deosebit de strânși (foarte condensați). Se adaugă colchicina otrăvitoare a fusului, care funcționează exact în această fază a ciclului celular, astfel încât cromozomii metafazici se acumulează. Apoi sunt izolate și colorate folosind metode speciale de colorare.
Cea mai comună este banda GTG, în care cromozomii sunt tratați cu tripsină, o enzimă digestivă și pigmentul Giemsa. Regiunile cu densitate deosebită și cele bogate în adenină și timină sunt prezentate întunecate.
Benzile G rezultate sunt caracteristice fiecărui cromozom și, în termeni simplificați, sunt considerate a fi regiunile cu mai puține gene. O imagine a cromozomilor colorați în acest mod este făcută la o mărire de o mie de ori și se creează o cariogramă cu ajutorul unui program de computer. În plus față de modelul benzii, dimensiunea cromozomului și poziția centromerului sunt utilizate pentru a ajuta la aranjarea cromozomilor în consecință. Dar există și alte metode de bandare care pot avea avantaje foarte diferite.
Recomandări din partea echipei editoriale
Pentru informații mai generale, consultați următoarele articole:
- Diviziunea nucleului celular
- Funcțiile nucleului celular
- Trisomia 21
- Boli genetice